
Three-Dimensional Aggregated Spheroid Model of Hepatocellular Carcinoma Using a 96-Pillar/Well Plate


Abstract
:1. Introduction
2. Results and Discussion
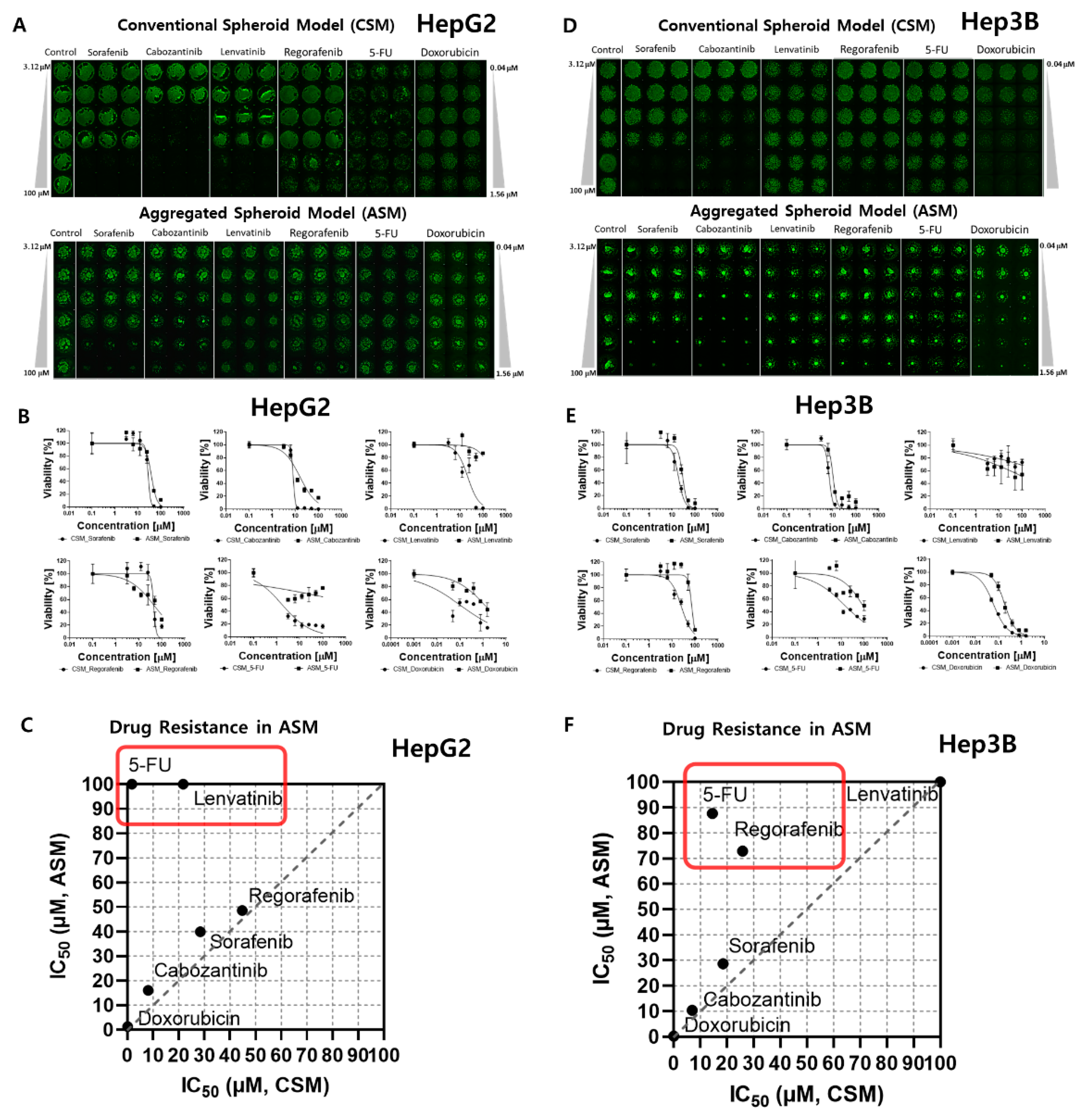
2.1. HTS of Six Anti-Cancer Drug Responses in Each 3D-Cell Model
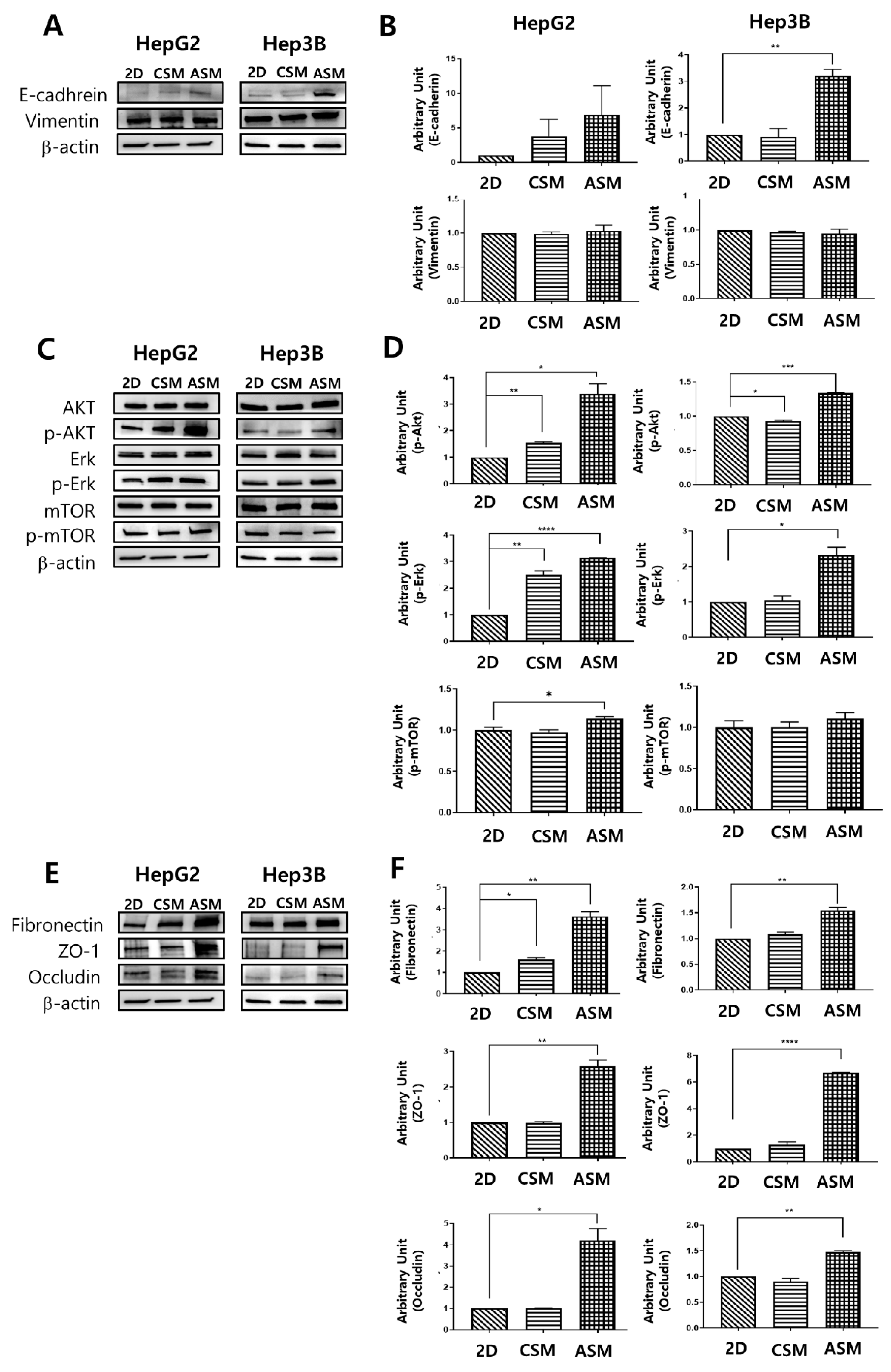
2.2. Western Blot Analysis of Epithelial Cell Markers, Cell Proliferation Receptor, and ECM/Cell Tight Junction Protein Expression
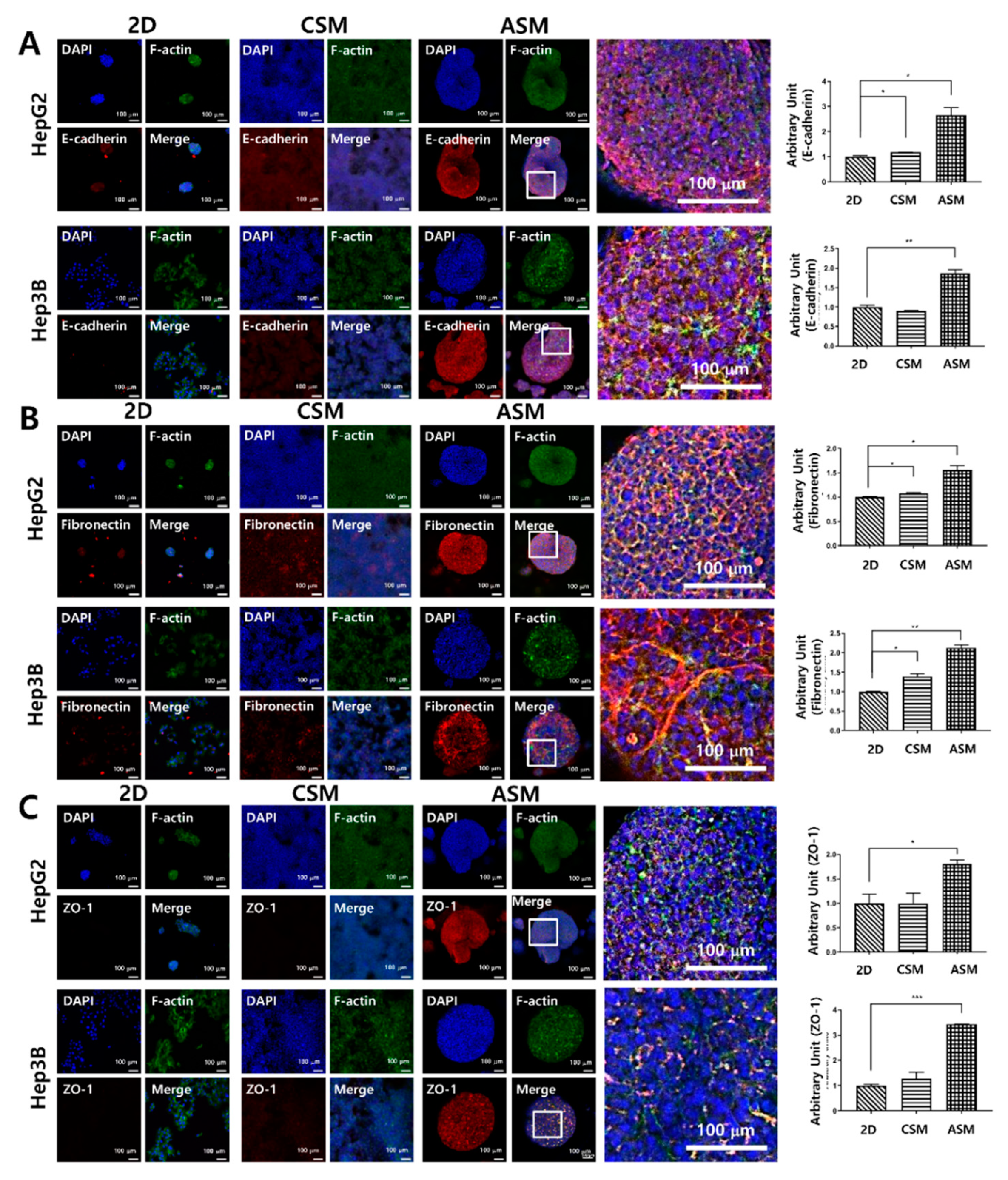
2.3. High Content Image Analysis of Epithelial Cell Markers, and ECM/Cell Tight Junction Protein Expression
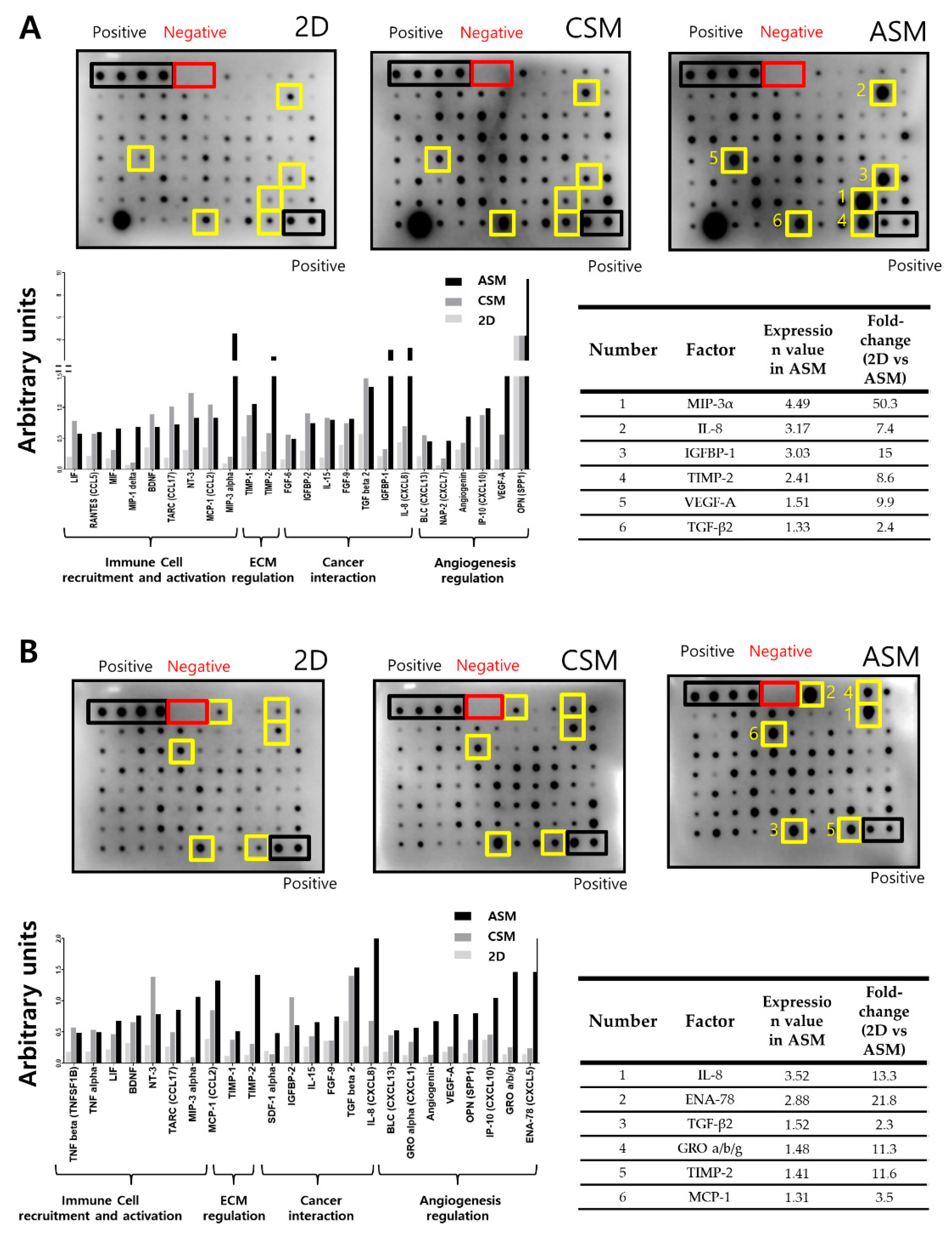
2.4. Cytokine Secretion Analysis Related to Cell Proliferation and Drug Resistance
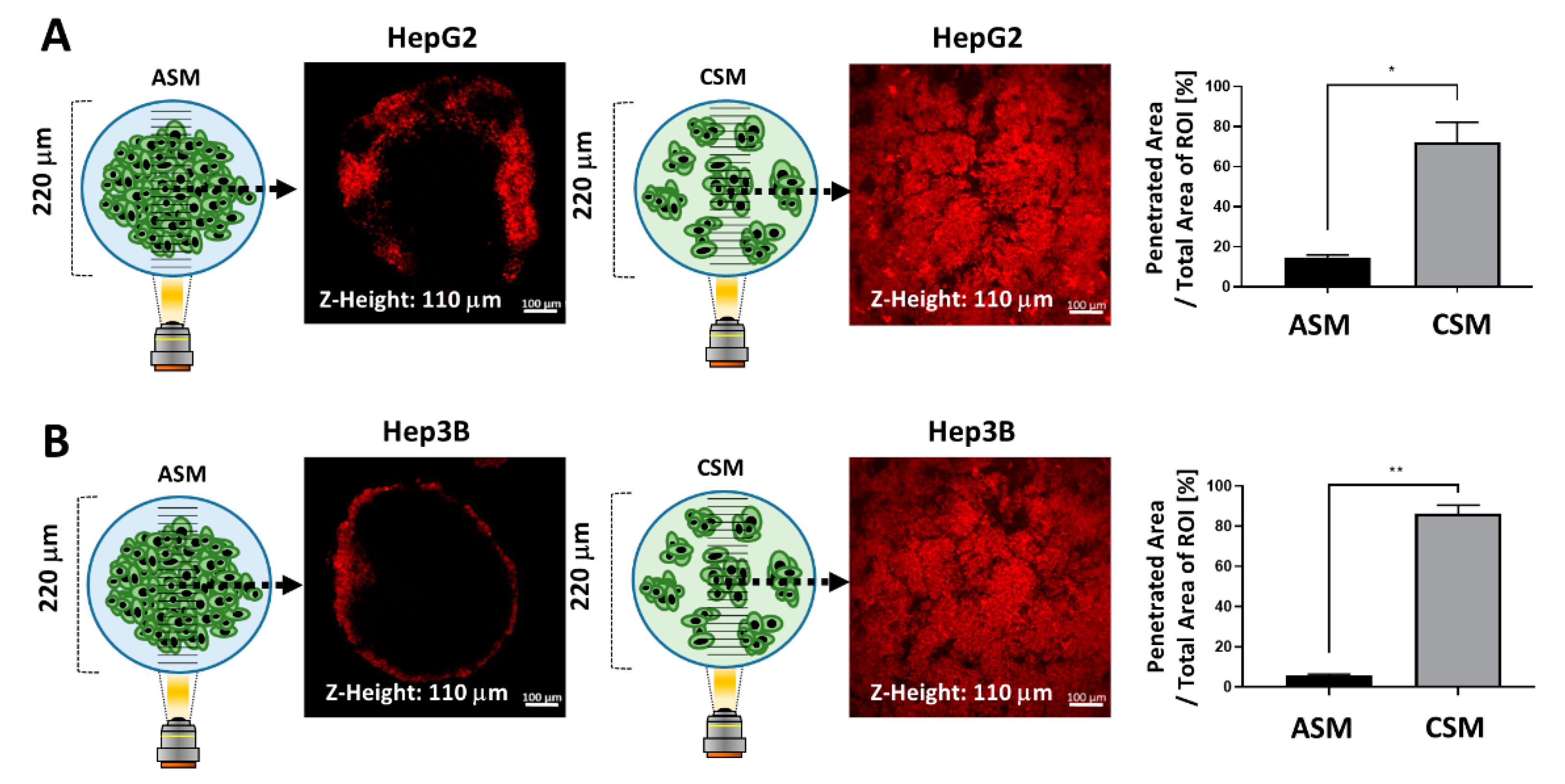
2.5. Drug penetration Assay for Validation of Drug Resistance in the ASM Using an Auto-Fluorescent Drug
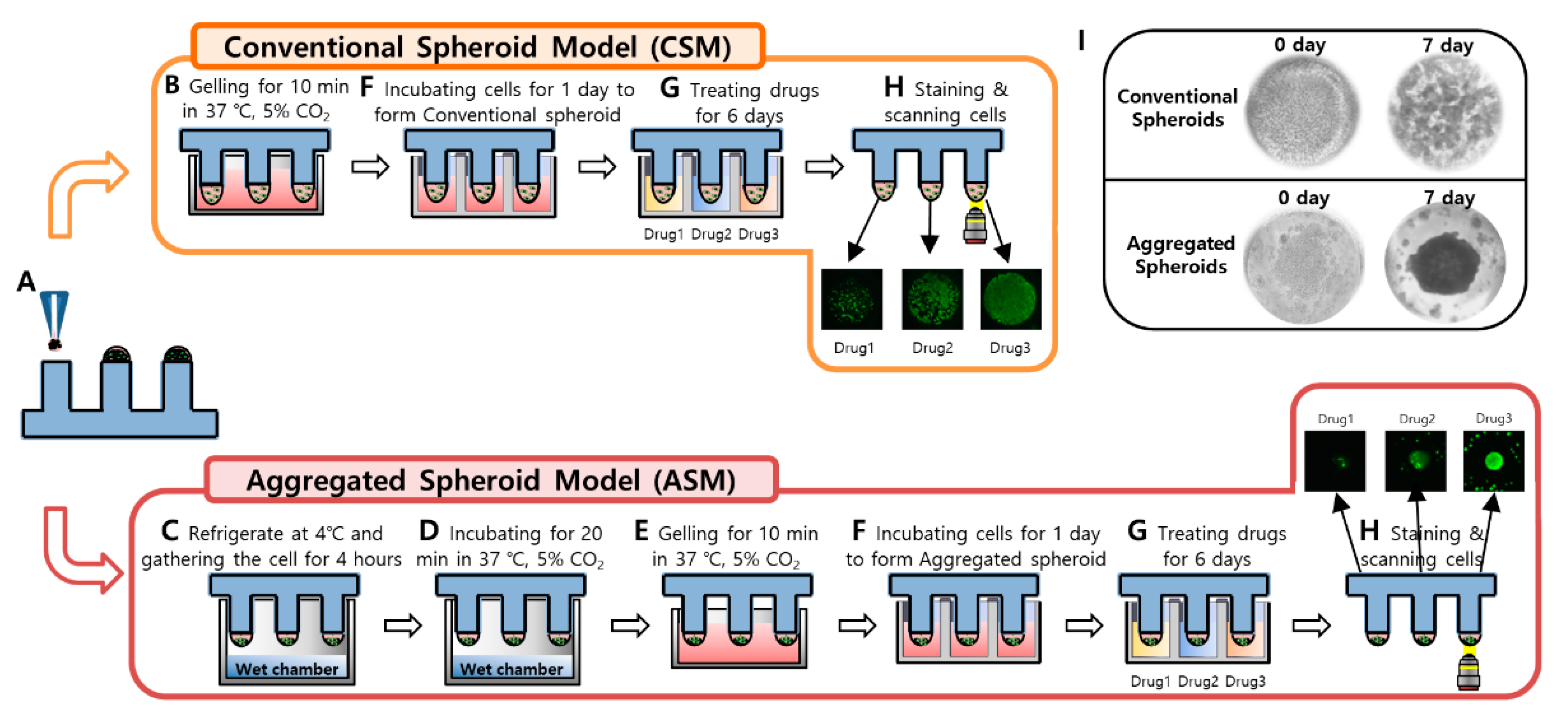
3. Materials and Methods
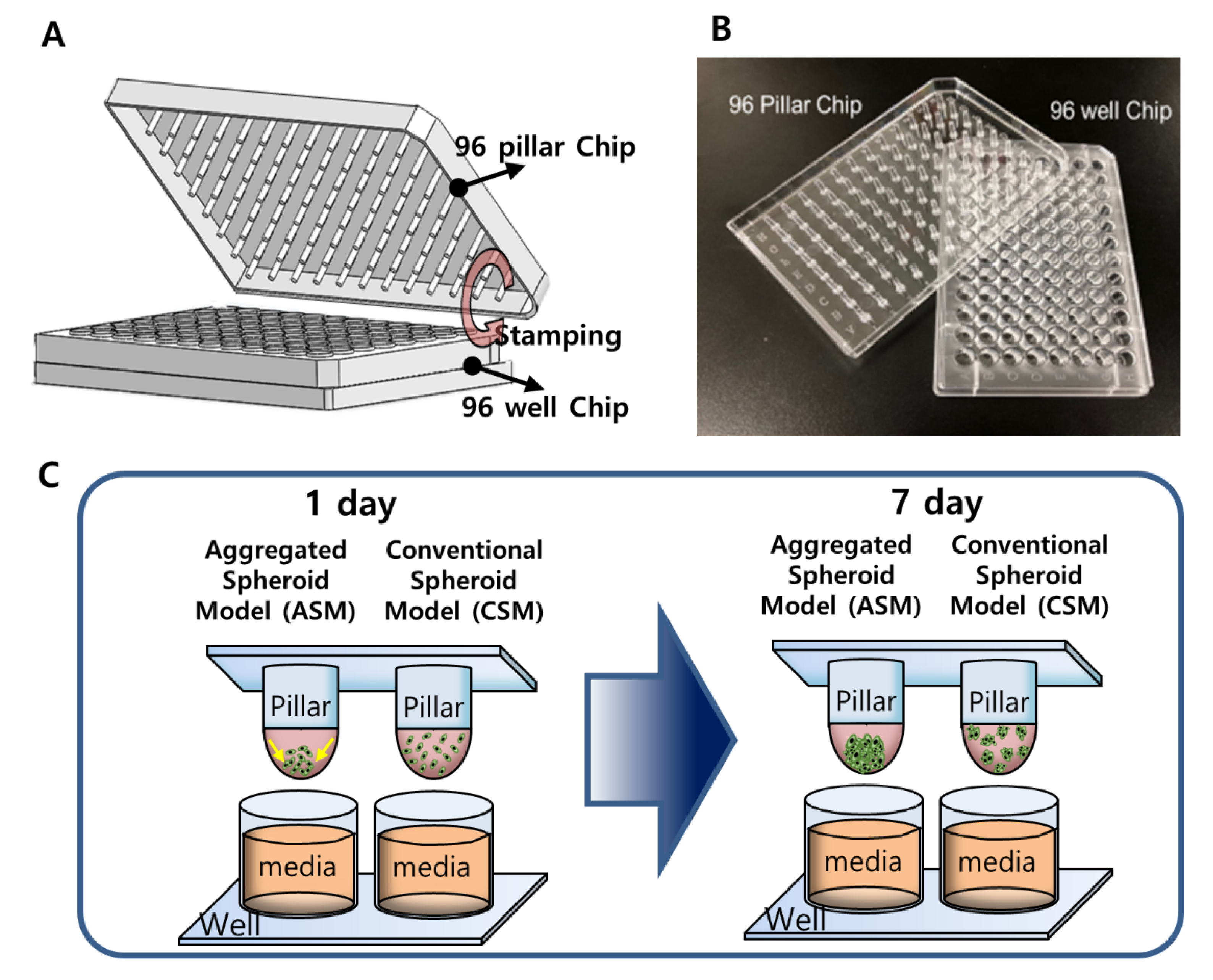
3.1. Preparation of the 96 Pillar/Well Plates
3.2. Experimental Procedure of High-Throughput Drug Screening
3.3. Experimental Procedure for Image Scanning and Data Analysis
3.4. Cell Culture
3.5. Drugs Preparation
3.6. 3D-Cell Viability Assay
3.7. Western Blot Analysis
3.8. High Content Image Analysis
3.9. Cytokine Assay
3.10. Drug Penetration Assay
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- El-Serag, H.B. Hepatocellular carcinoma. N. Engl. J. Med. 2011, 365, 1118–1127. [Google Scholar] [CrossRef]
- Lachenmayer, A.; Alsinet, C.; Chang, C.Y.; Llovet, J.M. Molecular approaches to treatment of hepatocellular carcinoma. Dig. Liver Dis. 2010, 42, 264–272. [Google Scholar] [CrossRef] [Green Version]
- Feitelson, M.A.; Sun, B.; Tufan, N.L.S.; Liu, J.; Pan, J.; Lian, Z. Genetic mechanisms of hepatocarcinogenesis. Oncogene 2002, 21, 2593–2604. [Google Scholar] [CrossRef] [Green Version]
- Edmondson, R.; Broglie, J.J.; Adcock, A.F.; Yang, L. Three-dimensional cell culture systems and their applications in drug discovery and cell-based biosensors. Assay Drug Dev. Technol. 2014, 12, 207–218. [Google Scholar] [CrossRef] [Green Version]
- Birgersdotter, A.; Sandberg, R.; Ernberg, I. Gene expression perturbation in vitro—a growing case for three-dimensional (3D) culture systems. Semin. Cancer Biol. 2005, 15, 405–412. [Google Scholar] [CrossRef]
- Weaver, V.M.; Petersen, O.W.; Wang, F.; Larabell, C.; Briand, P.; Damsky, C.; Bissell, M.J. Reversion of the malignant phenotype of human breast cells in three-dimensional culture and in vivo by integrin blocking antibodies. J. Cell Biol. 1997, 137, 231–245. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhadriraju, K.; Chen, C.S. Engineering cellular microenvironments to improve cell-based drug testing. Drug Discov. Today 2002, 7, 612–620. [Google Scholar] [CrossRef]
- Song, Y.; Kim, J.-S.; Kim, S.-H.; Park, Y.K.; Yu, E.; Kim, K.-H.; Seo, E.-J.; Oh, H.-B.; Lee, H.C.; Kim, K.M. Patient-derived multicellular tumor spheroids towards optimized treatment for patients with hepatocellular carcinoma. J. Exp. Clin. Cancer Res. 2018, 37, 109. [Google Scholar] [CrossRef]
- Shield, K.; Ackland, M.L.; Ahmed, N.; Rice, G.E. Multicellular spheroids in ovarian cancer metastases: Biology and pathology. Gynecol. Oncol. 2009, 113, 143–148. [Google Scholar] [CrossRef]
- Zietarska, M.; Maugard, C.M.; Filali-Mouhim, A.; Alam-Fahmy, M.; Tonin, P.N.; Provencher, D.M.; Mes-Masson, A.M. Molecular description of a 3D in vitro model for the study of epithelial ovarian cancer (EOC). Mol. Carcinog. 2007, 46, 872–885. [Google Scholar] [CrossRef]
- Le Roux, L.; Volgin, A.; Maxwell, D.; Ishihara, K.; Gelovani, J.; Schellingerhout, D. Optimizing imaging of three-dimensional multicellular tumor spheroids with fluorescent reporter proteins using confocal microscopy. Mol. Imaging 2008, 7, 214–221. [Google Scholar] [CrossRef] [Green Version]
- Tung, Y.-C.; Hsiao, A.Y.; Allen, S.G.; Torisawa, Y.-S.; Ho, M.; Takayama, S. High-throughput 3D spheroid culture and drug testing using a 384 hanging drop array. Analyst 2011, 136, 473–478. [Google Scholar] [CrossRef]
- Kang, J.; Lee, D.W.; Hwang, H.J.; Yeon, S.-E.; Lee, M.-Y.; Kuh, H.-J. Mini-pillar array for hydrogel-supported 3D culture and high-content histologic analysis of human tumor spheroids. Lab Chip 2016, 16, 2265–2276. [Google Scholar] [CrossRef]
- Lee, D.W.; Choi, Y.-S.; Seo, Y.J.; Lee, M.-Y.; Jeon, S.Y.; Ku, B.; Kim, S.; Yi, S.H.; Nam, D.-H. High-throughput screening (HTS) of anticancer drug efficacy on a micropillar/microwell chip platform. Anal. Chem. 2014, 86, 535–542. [Google Scholar] [CrossRef]
- Lee, D.W.; Doh, I.; Nam, D.-H. Unified 2D and 3D cell-based high-throughput screening platform using a micropillar/microwell chip. Sens. Actuators B 2016, 228, 523–528. [Google Scholar] [CrossRef]
- Lee, D.W.; Choi, Y.S.; Seo, Y.J.; Lee, M.Y.; Jeon, S.Y.; Ku, B.; Nam, D.H. High-Throughput, Miniaturized Clonogenic Analysis of a Limiting Dilution Assay on a Micropillar/Microwell Chip with Brain Tumor Cells. Small 2014, 10, 5098–5105. [Google Scholar] [CrossRef] [PubMed]
- Sebens, S.; Schafer, H. The tumor stroma as mediator of drug resistance-a potential target to improve cancer therapy. Curr. Pharm Biotechnol. 2012, 13, 2259–2272. [Google Scholar] [CrossRef]
- Fuchs, B.C.; Fujii, T.; Dorfman, J.D.; Goodwin, J.M.; Zhu, A.X.; Lanuti, M.; Tanabe, K.K. Epithelial-to-mesenchymal transition and integrin-linked kinase mediate sensitivity to epidermal growth factor receptor inhibition in human hepatoma cells. Cancer Res. 2008, 68, 2391–2399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breslin, S.; O’Driscoll, L. The relevance of using 3D cell cultures, in addition to 2D monolayer cultures, when evaluating breast cancer drug sensitivity and resistance. Oncotarget 2016, 7, 45745–45756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ye, Y.; Zhang, R.; Feng, H. Fibronectin promotes tumor cells growth and drugs resistance through a CDC42-YAP dependent signaling pathway in colorectal cancer. Cell Biol. Int. 2020, 9, 1840–1849. [Google Scholar] [CrossRef]
- Afasizheva, A.; Devine, A.; Tillman, H.; Fung, K.L.; Vieira, W.D.; Blehm, B.H.; Kotobuki, Y.; Busby, B.; Chen, E.I.; Tanner, K. Mitogen-activated protein kinase signaling causes malignant melanoma cells to differentially alter extracellular matrix biosynthesis to promote cell survival. BMC Cancer 2016, 16, 1–15. [Google Scholar] [CrossRef] [Green Version]
- Fedorenko, I.V.; Abel, E.V.; Koomen, J.M.; Fang, B.; Wood, E.R.; Chen, Y.A.; Fisher, K.J.; Iyengar, S.; Dahlman, K.B.; Wargo, J.A. Fibronectin induction abrogates the BRAF inhibitor response of BRAF V600E/PTEN-null melanoma cells. Oncogene 2016, 35, 1225–1235. [Google Scholar] [CrossRef] [Green Version]
- Rintoul, R.C.; Sethi, T. Extracellular matrix regulation of drug resistance in small-cell lung cancer. Clin. Sci. 2002, 102, 417–424. [Google Scholar] [CrossRef]
- Pankov, R.; Yamada, K.M. Fibronectin at a glance. J. Cell Sci. 2002, 115, 3861–3863. [Google Scholar] [CrossRef] [Green Version]
- Tunggal, J.A.; Helfrich, I.; Schmitz, A.; Schwarz, H.; Günzel, D.; Fromm, M.; Kemler, R.; Krieg, T.; Niessen, C.M. E-cadherin is essential for in vivo epidermal barrier function by regulating tight junctions. EMBO J. 2005, 24, 1146–1156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nebane, N.M.; Coric, T.; Whig, K.; McKellip, S.; Woods, L.; Sosa, M.; Sheppard, R.; Rasmussen, L.; Bjornsti, M.-A.; White, E.L. High-Throughput RNA Interference Screening: Tricks of the Trade. J. Lab. Autom. 2013, 18, 334–339. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HepG2 | Hep3B | ||||||
|---|---|---|---|---|---|---|---|
| Drug | Target | 2D-HTS | CSM | ASM | 2D-HTS | CSM | ASM |
| Sorafenib | VEGFR2-3, PDGFR-β, Raf-1, B-Raf | 39.06 | 28.45 | 39.89 | 20.1 | 18.56 | 28.65 |
| Cabozantinib | VEGFR2, c-Met | 5.65 | 8.11 | 16.08 | 4.79 | 6.99 | 10.35 |
| Lenvatinib | VEGFR1-3, FGFR1-4, PDGFRα | 34.94 | 21.77 | >100 | 1.443 | >100 | >100 |
| Regorafenib | VEGFR1-3, PDGFRβ, Kit, RET, Raf-1, B-RAF | 40.47 | 44.79 | 48.58 | 24.77 | 25.88 | 72.86 |
| 5-FU | DNA/RNA synthesis inhibitor | 1.49 | 1.78 | >100 | 7.71 | 14.5 | 87.6 |
| DOX | DNA damage, AMPK | 0.052 | 0.115 | 1.305 | 0.029 | 0.061 | 0.188 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, S.-Y.; Teng, Y.; Son, M.; Ku, B.; Hwang, H.J.; Tergaonkar, V.; Chow, P.K.-H.; Lee, D.W.; Nam, D.-H. Three-Dimensional Aggregated Spheroid Model of Hepatocellular Carcinoma Using a 96-Pillar/Well Plate. Molecules 2021, 26, 4949. https://doi.org/10.3390/molecules26164949
Lee S-Y, Teng Y, Son M, Ku B, Hwang HJ, Tergaonkar V, Chow PK-H, Lee DW, Nam D-H. Three-Dimensional Aggregated Spheroid Model of Hepatocellular Carcinoma Using a 96-Pillar/Well Plate. Molecules. 2021; 26(16):4949. https://doi.org/10.3390/molecules26164949
Chicago/Turabian StyleLee, Sang-Yun, Yvonne Teng, Miseol Son, Bosung Ku, Hyun Ju Hwang, Vinay Tergaonkar, Pierce Kah-Hoe Chow, Dong Woo Lee, and Do-Hyun Nam. 2021. "Three-Dimensional Aggregated Spheroid Model of Hepatocellular Carcinoma Using a 96-Pillar/Well Plate" Molecules 26, no. 16: 4949. https://doi.org/10.3390/molecules26164949